
2024年10月13日追記: イモリのゲノムについて
ハイギョという魚は、通常の魚と同様にエラ呼吸をすることに加え、肺呼吸することができる。我々は肺呼吸しかできないが、祖先をたどると、デボン紀頃、エラ呼吸だけをしていた魚から肺呼吸をする魚が出現した。
現生の魚で肺呼吸を残しているのはハイギョという魚とポリプテルスという魚だけである。ポリプテルスの魅力も筆舌に尽くしがたいが、今回はハイギョのゲノムについての新しい論文 (Schartl et al., 2024) が出たので、その解説を一般の人向けに書いてみようと思う。
ハイギョは大きく3つの地域に分布
ハイギョはシルル紀からデボン紀あたり(約4億年前)に出現し、そして今もなおその系統は絶えず続いている。原始的な肺など、古代の魚の形質を有していることから「生きた化石」などとも呼ばれている。私達四肢動物(両生類・爬虫類・哺乳類)と最も近縁な魚類であり、化石記録に基づくと、四肢動物とは 4.25 億年前に分岐したと言われている (Brownstein et al., 2023)。
場所的な起源は北アメリカ・ヨーロッパのあたりだと言われているが、かつては大陸が繋がっていた時期(パンゲア大陸など)もあり、おそらくはもっと幅広く生息していただろう。現に化石記録では少なくとも三畳紀(2.5-2.1 億年前)の地層からハイギョ類は南極大陸含む6大陸の全てで見つかっている (Eastman, 1991)。

しかし現在では南半球にしか生存しておらず、南アメリカ大陸、アフリカ大陸、オーストラリア大陸の3大陸にしか生存していない。それぞれの大陸に1科ずつあり、南アメリカ大陸、オーストラリア大陸のハイギョ(ミナミアメリカハイギョ、オーストラリアハイギョ)は1属1種である。アフリカハイギョは4種知られている。かつては70-100種いたと推測されており、ゴンドワナ大陸の分裂に伴って種分化が起きたと推定されている。最初に分裂したオーストラリアハイギョは他の2つの大陸のハイギョと遠縁である (Brownstein et al., 2023)。
実際、オーストラリアハイギョは他のハイギョよりも原始的な姿をしており、肉鰭 (分厚い胸鰭) を持っている。対して他の2大陸のハイギョはムチのようなヒレになっている。


ハイギョのゲノムはヒトの30倍大きい
さて、そんなハイギョだが、ゲノム(全ての遺伝情報を含むDNA)のサイズを見ると、他の脊椎動物とは比にならないくらい大きい。
2021年にハイギョ2種(アフリカハイギョとオーストラリアハイギョ)のゲノムが決定された (Bi et al., 2021; Meyer et al., 2021)。この論文では、アフリカハイギョ(Protopterus annectens) はゲノムサイズが約 40 Gb (ギガベース、DNAはATGCの4つの塩基からなるが、その塩基が40ギガ、つまり400億塩基)であり、オーストラリアハイギョは 37 Gb であるとされている。ヒトが約 3.2 Gb なので、10倍以上の大きさであることがわかる。
マウスやヒトなどをはじめとする「応用研究」に必須である生物や、家畜などのゲノムについてはこれまで多くの種が決定されてきていた。近年シーケンサ(DNA配列を決定する装置)の発展・低価格化によって多くのマイナーな種のゲノムも決定されてきたが、ゲノムが大きい生物は反復配列が多いことなどの理由でゲノム決定がされてこなかった。2021年の研究ではロングリードシーケンサというより長い配列を決定できる装置を駆使してゲノム決定を成し遂げた。
さて、今回2024年に新たに決定されたハイギョはミナミアメリカハイギョ(Lepidosiren paradoxa) である。このハイギョのゲノムサイズは 91 Gb あり、ヒトの約30倍の大きさである。これまで決定されてきたハイギョらよりも2倍以上大きい。
染色体比較により、初期四肢動物からどのように進化したかを見る
今回の論文では染色体同士を比較することで、初期四肢動物の染色体を推定している。その結果、アフリカハイギョ・ミナミアメリカハイギョでは一部の染色体が融合していることが観測された。一方で、オーストラリアハイギョではその初期四肢動物の染色体構造をよく保っているということが示された。このことは、オーストラリアハイギョのほうが他の大陸のハイギョより祖先の染色体構造を維持しているということを意味している。
なぜ染色体構造が変わってしまったのか、いろいろな理由はあるだろうが、著者らはトランスポゾンというゲノム内を動き回り、増殖する配列が原因ではないかと推測している。ハイギョの巨大なゲノムのほとんどはこのトランスポゾン配列によって占められており、この増えたトランスポゾンが染色体同士の融合や分裂、組み換えなどの不安定性に寄与しているのではないかとの見方だ。実際、トランスポゾンの配列の量で見ると、オーストラリアハイギョ (35 Gb) と比べ、アフリカハイギョ (40 Gb) とミナミアメリカハイギョ (85 Gb) の両者はより大きい。
また、全ゲノム重複というイベント後に増えた染色体のうちどちらかに偏って遺伝子が失われているということを示した。これを理解するために、まず全ゲノム重複について説明する。我々顎口類(顎のある脊椎動物)はこれまでに2回の全ゲノム重複を経験している。一度目はカンブリア紀に、自分自身の染色体が単純に二倍になることで、二度目はオルドビス紀に別々の種の交配によって異なる染色体が増えた(異質四倍体化)によって起きている。
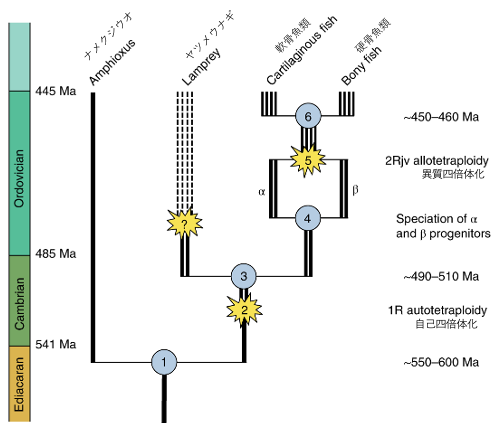
このとき、2回目の全ゲノム重複ではαとβという異なる由来の染色体が一つのゲノムに組み入れられたことになる。もちろん、遺伝子も一緒に。しかし大抵の場合、二つも同じ遺伝子が必要でない場合が多く、片方が失われる。異質四倍体化の場合、異なるエピジェネティクス、異なるトランスポゾンの活動などによって非対称の遺伝子消失や偽遺伝子化が起こることが知られている(Simakov et al., 2020)。
さて、それでは本題のハイギョゲノムの解析結果を見てみる。ガー(魚の一種)やオーストラリアハイギョではβとα両方の遺伝子が(アルファのほうが少ないものの)残っているのに対し、ミナミアメリカハイギョとアフリカハイギョではαの遺伝子が著しく偏って失われていることが判明した。このことは、オーストラリアハイギョのほうが他の2つの大陸のハイギョと比べて遺伝子の並び順(シンテニー)などが保存されているということを意味しているだろう。
ゲノム巨大化が起きた時期
ハイギョのゲノムは巨大であることは書いたとおりだが、ではいつ頃からその「巨大化」が起きたのだろうか?
100万年あたり、平均して 124 Mb の速度でゲノムが大きくなっていることがわかり、このことは現存するハイギョの共通祖先よりも前のハイギョの時代から巨大化が始まったことを意味している。そして、91 Gb という巨大なゲノムを生み出したミナミアメリカハイギョでは100万年あたり 3.71 Gb という速度でゲノムが大きくなり続けていることがわかった。これはこれまで知られているどの脊椎動物の中で最も速い速度である。
また、著者らは化石記録にも着目した。ゲノムが巨大になるとそれだけ核が大きくなり、必然的に細胞サイズも大きくなる。
以下の両生類の系統樹では細胞のサイズ(左)とゲノムサイズ(右)を示したものである。赤から青になるにかけてサイズが大きいことを示している。有尾両生類(イモリ・サンショウウオ)でもゲノムサイズが大きいことが知られており、これらの系統ではゲノムサイズの増加に伴って細胞サイズも大きくなることが知られている。
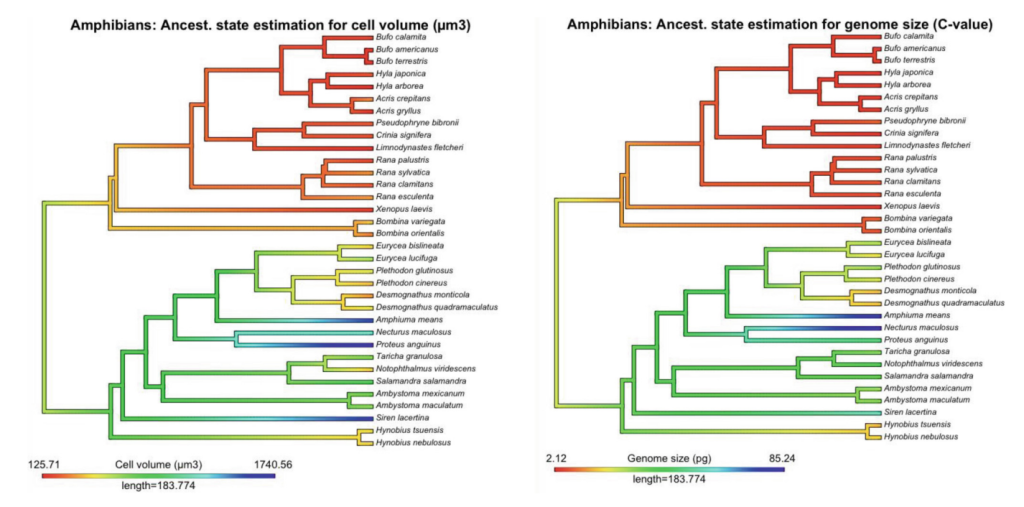
このことから、化石記録から細胞サイズを推定し(例えば骨細胞なんかを見る)、そこからゲノムサイズを推定することができる。結果としては、デボン紀の頃のハイギョ、Dipterus (ディプテルス) ではまだゲノムサイズが小さかったものの、その後の石炭紀の Gnathorhiza において少しずつ大きくなり、ペルム紀の Ceratodus (セラトダス / ケラトダス) でも大きくなり続けていた。そして現生ハイギョの共通祖先からオーストラリアハイギョが分岐後、オーストラリアハイギョの系統特異的にゲノムサイズが小さくなり、一方で2大陸のハイギョは大きくなり続けた、というシナリオが推定された。
何がゲノムを巨大にしたか?
巨大ゲノムの要因のほとんどは「トランスポゾン」や「繰り返し配列」である。これは以前のハイギョやアホロートル (ウーパールーパー) ゲノムのときに既に明らかになっていたことだ (Bi et al., 2021; Meyer et al., 2021; Nowoshilow et al., 2018)。
特にトランスポゾンは「Mobile element」と呼ばれるように、ゲノムの中を動き回る。ときにカット・アンド・ペースト、そしてコピー・アンド・ペーストによってゲノム中にその数を増やしていく。
そのトランスポゾンにも色々と種類があるのだが、ハイギョでは LINE と呼ばれる要素が多くを占めているのに対し、アホロートルでは LTR という要素が多くを占めている。このことはゲノムの巨大化という現象がハイギョとアホロートルでは独立に起きた可能性を示唆しているだろう。
さて、このトランスポゾンというものはゲノム上を飛び回るので基本的には宿主(そのゲノムをもつ生物)にとって有害なことが多い。なぜなら、機能遺伝子の中に挿入されようものならその遺伝子が破壊されかねないからだ。そのため、通常トランスポゾンの活性を抑制するような仕組みが備わっている。その一つに、PIWI-interacting RNAs (piRNAs) というものがある。この非コードRNAは生殖細胞系列においてトランスポゾンのコピー活性の抑制に関わることが知られている (Wang et al., 2023)。この piRNAs は、オーストラリアハイギョにおいて 28 bp の長さで発現のピークがきており、他の魚や両生類においてもその長さに発現のピークが来る。しかしアフリカハイギョとミナミアメリカハイギョではこの長さにピークは見られず、おそらく分解されているのではないかと考えられる。つまり、この piRNA のトランスポゾンに対する活性の低さがゲノム拡大の要因になっている可能性がある。
遺伝子とゲノムの進化
今回の論文では正の自然選択を受けた遺伝子を網羅的に探索している。この中には空気呼吸や肺呼吸に適した二重循環系、嗅覚などの遺伝子が含まれていた。また、面白いことに視床下部-下垂体-甲状腺系に関連した遺伝子も見つかっており、これはハイギョの幼生からの変態などに関連している可能性がある。
また、カルシウム代謝に関わる遺伝子も正の自然選択を受けていた。カルシウム代謝は魚類でエラが担っているが、四肢動物では副甲状腺が担う。そして副甲状腺は進化発生的にはエラ由来である(Okabe & Graham, 2004)。陸上適応に際して環境水中のカルシウムが取り込みにくくなることから、血中カルシウム濃度の調節のためにそうした遺伝子に選択が働いた可能性がある。
まとめと感想
本論文では、91 Gb という超巨大なハイギョのゲノムを決定した。これは偉業である。2021年のハイギョ2種とポリプテルスゲノムの論文が私の中でここ10年、期待を上回るワクワクした瞬間だったが、またこれが2024年になっても味わえるとは思わなかった。
ハイギョは最も両生類に系統的に近い現生魚類であることから、そのゲノム進化を理解することは陸上適応を理解するうえで重要である。
今回、以前からも示されてはいたが、改めてトランスポゾンのゲノム中の伝播がゲノム拡大に寄与しており、そして今もなおゲノムは拡大中であることを示した。まだまだ大きくなれるね。
こんなにゲノムが繰り返し配列だらけになってしまうと、普通なら染色体同士の組み換えなどにより、どんどん祖先型の染色体構造が失われてしまう。しかしハイギョでは大きな構造は保たれていたという特殊な一面も明らかになった。
今回記事には含めていないが、論文中ではヒレの発生についても触れている。オーストラリアハイギョでは「肉鰭類」の名に恥じぬ立派なヒレを持っており、およそ1億年以上に渡ってその形態はほとんど変化していないようである。一方でアフリカハイギョとミナミアメリカハイギョではムチのようなヒレとなってしまっている。論文中では Shh (ソニック・ヘッジホッグ) シグナルに関わる ZRS の変異が関わっていることを示している。もし四肢発生に興味がある方はぜひ読んでみて欲しい。
これで三大陸のハイギョゲノムが出揃った。本論文タイトルは「The genomes of all lungfish inform on genome expansion and tetrapod evolution」、つまり「全ハイギョのゲノムから、ゲノムの拡大と四肢動物の進化を解明する」というものであるが、まだゲノムが決定していないハイギョも存在する。プロトプテルス属ではアネクテンス (Protopterus annectens) しか決定されていない。
また、独立に巨大化が起きたものの、有尾両生類 (サンショウウオ、イモリ) のゲノムも巨大である。現在公開されているゲノムはアホロートル(ウーパールーパー)とイベリアトゲイモリだけである。他にはホライモリのゲノムプロジェクトも進んでいるようだが、イベリアトゲイモリと同じくまだ論文が雑誌に掲載されるには至っていない。今後の巨大ゲノム界隈の発展を祈っている。
2024年10月13日追記
この記事を書いた数日後、NCBIのゲノム情報を漁っていたところ、
- Triturus cristatus (warty newt、ホクオウクシイモリ)
- Calotriton arnoldi (Montseny brook newt)
- Lissotriton vulgaris (common newt、スベイモリ)
- Lissotriton helveticus (palmate newt、ヒラユビイモリ)
のゲノムが公開されていた。そのうち Calotriton arnoldi については既にゲノム論文が公開されているので、利用可能な状態だ (Talavera et al., 2024)。
参考文献
- Schartl, M., Woltering, J. M., Irisarri, I., Du, K., Kneitz, S., Pippel, M., … & Meyer, A. (2024). The genomes of all lungfish inform on genome expansion and tetrapod evolution. Nature, 1-8. https://doi.org/10.1038/s41586-024-07830-1
- Brownstein, C. D., Harrington, R. C., & Near, T. J. (2023). The biogeography of extant lungfishes traces the breakup of Gondwana. Journal of Biogeography, 50(7), 1191-1198.
- Eastman, J. T. (1991). The fossil and modern fish faunas of Antarctica: evolution and diversity. In Biology of Antarctic Fish (pp. 116-130). Berlin, Heidelberg: Springer Berlin Heidelberg.
- Bi, X., Wang, K., Yang, L., Pan, H., Jiang, H., Wei, Q., … & Zhang, G. (2021). Tracing the genetic footprints of vertebrate landing in non-teleost ray-finned fishes. Cell, 184(5), 1377-1391.
- Meyer, A., Schloissnig, S., Franchini, P., Du, K., Woltering, J. M., Irisarri, I., … & Schartl, M. (2021). Giant lungfish genome elucidates the conquest of land by vertebrates. Nature, 590(7845), 284-289.
- Simakov, O., Marlétaz, F., Yue, J. X., O’Connell, B., Jenkins, J., Brandt, A., … & Rokhsar, D. S. (2020). Deeply conserved synteny resolves early events in vertebrate evolution. Nature ecology & evolution, 4(6), 820-830.
- Wilkins, J. S. (2023). Species, God, and Dominion. In Speciesism in Biology and Culture: How Human Exceptionalism is Pushing Planetary Boundaries (pp. 95-110). Cham: Springer International Publishing.
- Nowoshilow, S., Schloissnig, S., Fei, J. F., Dahl, A., Pang, A. W., Pippel, M., … & Myers, E. W. (2018). The axolotl genome and the evolution of key tissue formation regulators. Nature, 554(7690), 50-55.
- Wang, J., Yuan, L., Tang, J., Liu, J., Sun, C., Itgen, M. W., … & Mueller, R. L. (2023). Transposable element and host silencing activity in gigantic genomes. Frontiers in Cell and Developmental Biology, 11, 1124374.
- Okabe, M., & Graham, A. (2004). The origin of the parathyroid gland. Proceedings of the national academy of sciences, 101(51), 17716-17719.
- Talavera, A., Palmada-Flores, M., Burriel-Carranza, B., Valbuena-Ureña, E., Mochales-Riaño, G., Adams, D. C., … & Carranza, S. (2024). Genomic insights into the Montseny brook newt (Calotriton arnoldi), a Critically Endangered glacial relict. Iscience, 27(1).




![【Qiskit】マルチオミクス解析を量子機械学習でやる①[環境構築・基礎]](https://kimbio.info/wp-content/uploads/2024/05/2203027-100x100.jpg)


はじめまして。突然のコメント失礼いたします。
最近疑問に思う事があり、インターネットで調べていたところ当サイトを見つけ、拝見させていただきました。
自分は酪農家で、数学も生物も無知な素人です。
自分はコラッツ予想という数学の問題について、この数式は生物の雌雄を決定する上で、雌雄割合を50%を保つ為に作用する法則では無いかと仮定しています。この仮説はどこかに存在していて、検証を試みる方が居るのか生物学に知見のある方にお聞きしたいと思いコメントさせて頂きました。
はじめまして、コメントありがとうございます。
進化生物学の関係で、コラッツ予想と雌雄比について絡めた考察というのはあまりされていないと思います。
論文を探すと一本、人工生命と絡めたものが見つかりました。ただ性比ではなく、個体群の数を見ているようです。
通説的に、雌雄比が1:1に保たれるのはフィッシャーの原理というものに従うとされています。
昔書いた記事にだいたいの解説が載っています。
進化の上で、性比の偏りが生じるとそれに反するように繁殖するほうが進化的に有利(子孫を多く残せる)ようになるため、結局1:1に収束するというものです(ただし必ずしも1:1ではなく、例外も多く見つかっています)。
子孫を残すということと、コラッツ問題における「操作」は似たものに感じられるかもしれませんが、生物では「より多くの子孫を残すことが大事」という根底が存在するので両者に科学的には関係があるとは言えないと思われます。
ただ、有向グラフを見てみると生物における系統樹にも見えますね。興味深い着眼点だと思います。